EVALUACIÓN DEL EFECTO DE FEROMONA DE CAMINO Y COMPORTAMIENTO AGONÍSTICO ENTRE COLONIAS DE Nasutitermes sp.
Las feromonas representan el principal tipo de comunicación química entre insectos, pudiendo mediar en la toma de decisiones sobre el item alimenticio a explotar y en las respuestas de agresión entre individuos intercoloniales. Por ello se tuvo por objetivo evaluar el efecto de la feromona de camino y el comportamiento agonístico entre colonias de Nasutitermes sp. Se utilizaron láminas de acetato, sobre las cuales se colocaron dos líneas paralelas, con extracto de abdómenes y hexano, en un extremo se colocó una capsula perforada con 7 obreras y 3 soldados de Nasutitermes sp. Se cronometro el tiempo de salida del primer individuo, el número y la casta de los individuos que llegaron al final la línea en 10 minutos. Se marcaron con corrector de bolígrafo, los abdómenes de 6 obreras de las colonias A y B. Se realizaron encuentros entre los miembros de una misma colonia y entre miembros de ambas colonias. Se contabilizaron el número de comportamientos neutrales, de alarma o agresión, por diez periodos de 30 segundos cada uno. Los resultados se evaluaron con una prueba paramétrica de ANOVA. Se obtuvo que no existieron diferencias significativas en la captación de hexano o extracto de abdómenes, sin embargo, el número de individuos que culminan el recorrido es mayor con el estímulo de feromonas. Además se observaron comportamientos agonísticos entres las colonias A y B. No hubo homogeneidad de varianza entre los tipos de interacción neutral, alarma y agresión, pero si hubo un marcado ataque ante individuos que no pertenecían a la misma colonia.
INTRODUCCIÓN
Las feromonas son señales químicas que median la comunicación entre miembros de una misma especie. Estos compuestos son producidos dentro del animal y luego liberados al ambiente. En muchos animales, estas sustancias transmiten información relacionada al estatus social como el género o dominancia y evocan comportamientos que son típicamente estereotipados y no se modifican con la experiencia. (Randall et al., 2002).
Las feromonas de camino son importantes principalmente en insectos sociales (abejas, avispas, hormigas y termitas) y subsociales. La feromona de camino es un rastro que dejan los forrajeros sobre el sustrato entre el nido y el alimento, y está conformado por componentes volátiles y componentes persistentes. En insectos sociales estas sustancias también se encuentran relacionadas al sistema de toma de decisiones sobre el ítem alimenticio a explotar (Hill et al., 2006). Por otra parte, la agresión se encuentra ligada al reconocimiento de los individuos emparentados pertenecientes a la misma colonia. En esta acción intervienen diversas señales como componentes químicos de la cutícula, estímulos táctiles, exudados glandulares y patrones de comportamiento propios de cada especie (Espíndola, 2011). El reconocimiento y el espaciamiento de la colonia son regulados por los hidrocarburos cuticulares. Estos compuestos de cera se producen sobre la cutícula exterior de las termitas y se esparcen a través de la colonia. Las termitas también pueden comunicar el peligro mediante golpes en la cabeza de soldados, el toque de cabeza en las galerías sirve para alertar a sus compañeros de nido (Resh & Cardé, 2009).
Es por ello que se tuvo por objetivo evaluar el efecto de la feromona de camino y el comportamiento agonístico entre colonias de Nasutitermes sp.
MATERIALES Y MÉTODOS
1.1 Orientación química en Nasutitermes sp
Sobre una hoja blanca con dos líneas paralelas, de 25 cm cada una y separadas entre sí por 18 cm, se colocó una lámina de acetato. Encima de cada línea se aplicó extracto de abdómenes y en un extremo una cápsula perforada que contenía 7 obreras y 3 soldados de Nasutitermes sp. Se cronometro el tiempo que tardo el primer individuo en comenzar a caminar sobre la línea; también se contabilizaron el número y la casta de los individuos que llegaron al final la línea en 10 minutos de registro. Este ensayo se repitió 5 veces, cambiando la lámina de acetato en cada uno de ellos. Se siguió la misma metodología para la evaluación del efecto del hexano. Los datos obtenidos fueron evaluados con una prueba de ANOVA.
1.2 Reconocimiento químico de individuos de diferentes colonias.
Se marcaron con corrector de bolígrafo, los abdómenes de 6 obreras de Nasutitermes sp pertenecientes a la colonia A y 6 obreras de la colonia B. Cada grupo fue colocado en una cápsula de Petri por 15 minutos, transcurrido ese tiempo, se realizaron cuatro encuentros: entre los miembros de colonia A, entre los miembros de la colonia B, entre los miembros teñidos de la colonia A y los no teñidos de la colonia B y viceversa. En dichos encuentros se observaron bajo una lupa y se contabilizaron el número de comportamientos asociados a cada tipo de encuentro (neutral, alarma o agresión) por diez periodos de 30 segundos cada uno. Se calculó el promedio del número de veces que se repite cada comportamiento durante los 10 conteos realizados y se realizó una prueba de Kruskall-Wallis, para determinar si habían diferencias entre los tipos de interacción por parte de las termitas ante los encuentra intra e intercoloniales.
Resultados y Discusión
1.1 Orientación química en Nasutitermes sp
En las tablas I y II se presenta el tiempo de salida del primer individuo de la colonia A, así como el número de individuos que llegan al final del recorrido con Hexano y extracto de abdómenes. En esta tabla se observa que los soldados fueron los primeros en salir ante los diferentes estímulos químicos, exploraron el camino y luego se retiraron para dar paso a las obreras, de allí que todas las termitas que llegaron al final del recorrido fueran de esta casta. El comportamiento observado concuerda con lo descrito por Arab & Issa (2000), en el que reportaron que la búsqueda es iniciada por pequeños grupos de soldados de 2-5 individuos, que salen del nido en todas direcciones. Cuando uno de estos soldados encuentra una fuente de alimento, comienza a presionar su abdomen contra el sustrato; este comportamiento está asociado a la deposición de feromona de camino (Stuart, 1969). Esta feromona es segregada por la glándula esternal que se encuentra entre el cuarto y quinto esternito abdominal, marcando el camino de orientación para las obreras. (Noirot y Noirot-Timothèe 1965; Stuart, 1969). Un comportamiento de forrajeo similar, iniciado por los soldados, fue observado para N. costalis por Traniello (1981). Sin embargo, la casta que comienza el reclutamiento puede ser diferente en otras las especies (Pasteels, 1965).
En estas tablas también se observa que, en promedio, el tiempo de salida del primer individuo es menor con el rastro de feromonas y el número de ejemplares de Nasutitrermes sp que llega al final de la marca es mayor con este estimulo químico.
Tabla I Tiempo de salida del primer individuo de la colonia A que llegan al final del recorrido.
| Replica | Hexano | Extracto | Casta |
|---|---|---|---|
| 1 | 5 | 27 | Soldado |
| 2 | 30 | 7 | Soldado |
| 3 | 18 | 9 | Soldado |
| 4 | 52 | 10 | Obrera |
| 5 | 76 | 9 | Obrera |
| Promedio | 36,2 | 12,4 | - |
Tabla II Número de terminas de la colonia A que llegan al final del recorrido
| Replica | Hexano | Extracto | Casta |
|---|---|---|---|
| 1 | 2 | 6 | Obrera |
| 2 | 1 | 10 | Obrera |
| 3 | 0 | 5 | Obrera |
| 4 | 1 | 5 | Obrera |
| 5 | 2 | 6 | Obrera |
| Promedio | 1,2 | 6,2 | - |
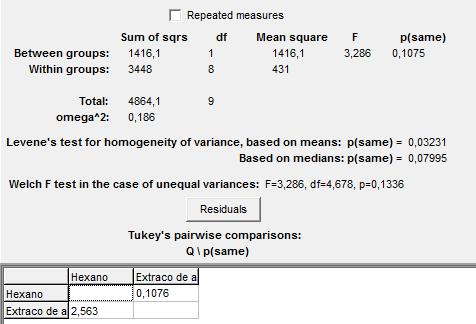
Para evaluar la homogeneidad de varianzas entre el tiempo de salida de los primeros individuos de la colonia A, con hexano y extracto de abdómenes; se aplicó una prueba de ANOVA, previa comprobación de la distribución normal de los datos, los resultados se ilustran en la figura 1. En esta prueba estadística, se tuvo por hipótesis nula la igualdad de las varianzas y por hipótesis alternativa su diferencia. Con un p-valor de 0,1075 y un 95% de confianza se mantiene la hipótesis nula; por lo tanto no existen diferencias signifi-cativas entre los tiempos de salida de los primeros individuos de la colina A, ante ambos es-tímulos químicos.
Figura 1: Prueba de ANOVA para el tiempo de salida del primer individuo de la colonia A, con hexano y extracto de abdómenes.
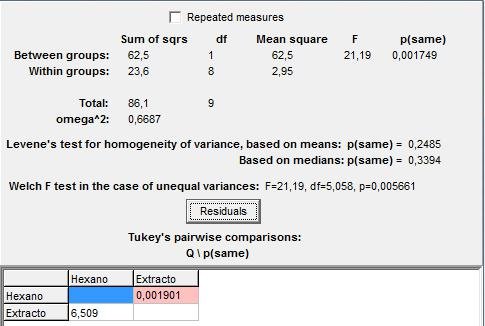
De forma homóloga también se evaluó con una prueba de ANOVA la homogeneidad de varianzas en el número de termitas que llegan al final del camino de he-xano y extracto de abdómenes de la colonia A. Los resultados se presentan en la figura 2. Con un p-valor de 0,0017 se mantiene la diferencia en la homogeneidad de las varianzas y por tanto se afirma que existen diferencias significativas entre el número de individuos de Nasutitrermes sp que llegan hasta el final del recorrido, siendo mayor el número de termitas que siguen el extracto de abdómenes o feromona de camino.
Figura 2: Prueba de ANOVA para el numero de individuo de la colonia A, que llevan al final de recorrido con hexano y extracto de abdómenes.
En las tablas III y IV, se presentan las respuestas de individuos de Nasutitermes sp en presencia de extracto de abdómenes y de hexano. En ella se ob-serva que los individuos de la colonia B, tardan 30,2 segundos más, en seguir el camino con extracto de abdómenes que el de hexano; además los primeros individuos en salir fueron en su mayoría termitas obreras. Estos resultados no concuerdan con lo reportado en estudios previos y difiere a su vez, con los resultados obtenidos en la colonia A; lo cual, índico que el forrajero, puede variar entre colonias de individuos de la misma especie. En cuanto al núme-ro de termitas que culminan el recorrido se obtuvieron resultados similares a los reportados en la tabla I, siendo el extracto de abdómenes el estímulo químico que atrae a más indivi-duos obreros de la especie de interés.
Tabla III Tiempo de salida del primer individuo de la colonia B que llegan al final del recorrido.
| Replica | Hexano | Extracto | Casta |
|---|---|---|---|
| 1 | 49 | 135 | Obrera |
| 2 | 3 | 39 | Soldado |
| 3 | 62 | 24 | Obrera |
| 4 | 9 | 7 | Obrera |
| 5 | 10 | 79 | Obrera |
| Promedio | 26,6 | 56,8 | - |
Tabla IV Número de terminas de la colonia B que llegan al final del recorrido
| Replica | Hexano | Extracto | Casta |
|---|---|---|---|
| 1 | 2 | 10 | Obrera |
| 2 | 1 | 4 | Obrera |
| 3 | 3 | 6 | Obrera |
| 4 | 4 | 4 | Obrera |
| 5 | 2 | 5 | Obrera |
| Promedio | 2,4 | 5,8 | - |
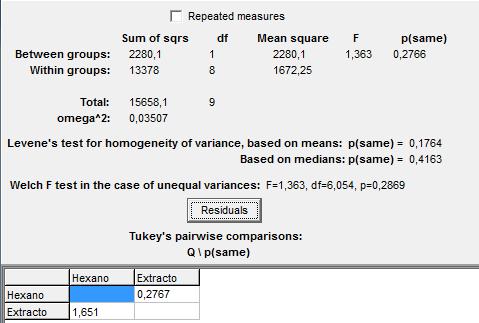
En la figura 3 se presentan los resultados de una prueba de ANOVA aplicada para evaluar si existen diferencias significativas entre el tiempo de salida de los primeros individuos de la colonia B de la especie de prueba. En base al p-valor (0,2766) se mantiene que no hay diferencias entre los tiempos de captación del estímulo químico con hexano y extracto de feromonas de camino.
Figura 3: Prueba de ANOVA para el tiempo de salida de los primeros individuos de la colonia B, con hexano y extracto de abdómenes.
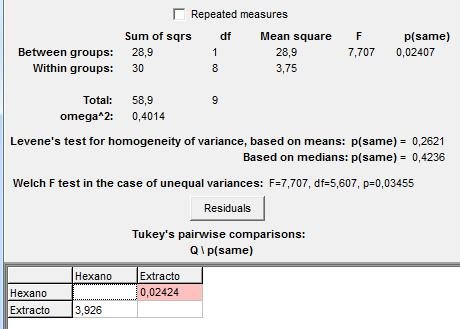
En la figura 4 se presentan los resultados de la prueba de ANOVA aplicada al número de termitas de la colonia B, que culminan el recorrido de hexano y feromonas. El p-valor de esta prueba fue de 0,02407, por ser menor a 0,05 se mantiene la hipótesis nula que plantea que las varianzas son diferentes y por tanto existen diferencias altamente significativas para afirmar que el número de termitas que llegan al final del recorrido de feromonas es mayor al número de individuos que culminan el recorrido de hexano.
Figura 4: ANOVA , para el numero de termitas de la colonia B que culminan el recorrido con hexano y extracto de abdómenes.
1.2 Reconocimiento químico de individuos de diferentes colonias
Se pudo observar también el comportamiento que hubo en las colonias tanto inter como intra, desempeñándose un papel de comportamiento neutral y de alarma en las termitas pertenecientes a la misma colonia (tanto colonias A como B), en cambio al detallar el comportamiento de termitas provenientes de diferentes colonias hubo interacción neutral, alarma y agresión hacia las “intrusas” en el encuentro A teñidas/B, y solo alarma y ataque en el encuentro A/B teñidas. Esto se debe a que el papel de las feromonas de alarma, producidas por diversas glándulas en insectos sociales o circunstancialmente agrupados, es sutil y siempre adaptado a los eventos presentes, siguiendo de esta forma ciertas pautas de comportamiento que consisten en una atracción primaria hacia la fuente de emisiones, seguida de un proceso de defensa y dispersión (Guerrero, 1988).
Por ejemplo, las termitas australianas Drepanotermes rubriceps exhiben una serie de movimientos oscilatorios cuando son molestadas, a la vez que emiten una feromona de alarma que induce sobre las demás, repetidos intentos de morder al agresor, lo que coincide perfectamente con el comportamiento observado en el laboratorio con organismos de diferentes dominio (Moore, 1968). Al realizar una prueba paramétrica de ANOVA, se observó que, al comparar la data por tipo de interacción entre las termitas de la misma y distintas colonias para determinar si hay homogeneidad de varianza en su comportamiento se realizo el siguiente planteamiento de hipótesis:
H0: Existe homogeneidad de varianza
H1: No existe homogeneidad de varianza.
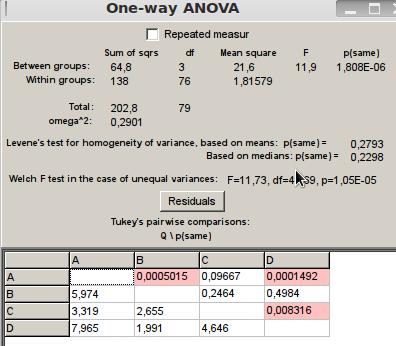
Se evidencia que la hipótesis nula se rechaza con un p(same) de 0,000001808<0,05 por lo que no hay homogeneidad de varianza, al tomar como posteriori la prueba de Tukey se identifica que en la interacción de neutralidad bien sea por anteneo o palpación de las termitas del tratamiento A fue diferente a las termitas del tratamiento B, pero iguales al tratamiento C. Este resultado coincide con el esperado, ya que las termitas del tratamiento A y C pertenecían a la misma colonia por lo que se confiaba en que presentaran la misma conducta. Así mismo las colonias B y D exhibieron el mismo comportamiento por el mismo razonamiento explicado anteriormente (Figura 5).
Figura 5: ANOVA para interacción del tipo neutral.
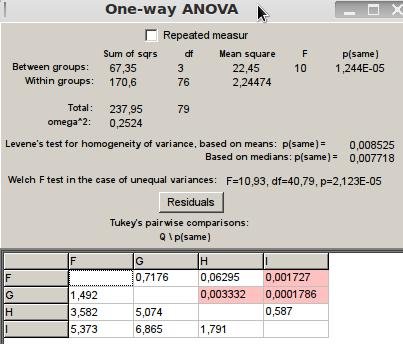
Al realizar el mismo procedimiento para la interacción de alarma o evitación y persecución, se rechaza la hipótesis nula con un p(same) de 0,00001244<0,05 por lo que no hay homogeneidad de varianza, al realizar la prueba posteriori se evidencia que los comportamientos de las termitas en los tratamientos F, G y H son iguales, y al mismo tiempo diferentes a I, esto se debe a que en todos los tratamientos hubo una llamada de alerta entre los organismos siendo menos evidente en el último (Figura 6).
Figura 6: ANOVA para interacción del tipo alarma.
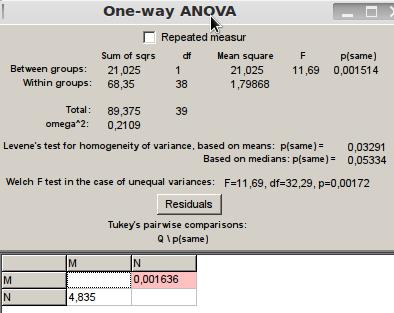
Finalmente para la interacción de agresión en la que se demostró lucha, pellizcos y mordidas entre las termitas, solo se hizo uso de los datos en los tratamientos 3 y 4, ya que en los tratamientos 1 y 2 no hubo señal de ataque debido a que las termitas a evaluar pertenecían a la misma colonia respectivamente, por lo que al haber reconocimiento entre ellas no se presentaba disputa alguna. Al realizar la prueba de ANOVA se rechazó la hipótesis nula con un p(same) de 0,001514<0,05 por lo que no hubo homogeneidad de varianza, siendo el comportamiento entre las termitas provenientes de distintas colonias diferentes en cuanto a nivel de agresión, pero manteniendo el hecho que al reconocer que algunos de los individuos presentes en el tratamiento no pertenecían a la misma sociedad, ingresaban en modo defensa atacando a las "invasoras”, hecho ocurrido en los dos tratamientos a detallar (3 y 4) (Figura 7).
Figura 7. ANOVA para interacción del tipo agresión.
CONCLUSIONES
- El forrajero varió entre las colonias A y B de Nasutitrermes sp, siendo los soldados, de la colonia A, los primeros en explorar el recorrido; mientras que en la colonia B fueron las obreras las primeras en salir.
- No existen diferencias significativas entre el tiempo de salida de los primeros individuos, de ambas colonias, con el hexano y el extracto de abdómenes.
- Con un 95% de confianza se afirma que el número de individuos de las colonias A y B de Nasutitrermes sp. que llegan al final del recorrido de extracto de abdómenes (feromona de camino) es mayor al que culminan el recorrido de hexano.
- No hubo homogeneidad de varianza en ninguna de las prueba paramétricas evaluadas ante las interacciones de neutralidad, alarma y agresión.
- La interacción de alarma fue notable en todos los tratamientos debido a que por medio de este comportamiento se determina si el individuo pertenece a la colonia o representa una amenaza.
- Fue notable la agresión ante la presencia de termitas provenientes de distintas colonias, comportamiento que se atribuye a la presencia de feromonas distintas en cada termitero que permite diferenciar organismos ajenos a la sociedad.
REFERENCIAS BIBLIOGRÁFICAS
- Arab A, Issa S. (2000). Breves observaciones sobre el comportamiento de forrajeo de dos especies de termitas (Termitidae: Nasutitermitinae) bajo condiciones de laboratorio. Bol. Entomol. Venez. 15(1):93-95.
- Espíndola, V. (2011) Comportamiento Agonístico de Obreras de Termitidae (Isoptera: Nasutitermitinae y Apicotermitinae). Zoología de invertebrados. Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE). Corrientes, Argentina.
- Guerrero, A. (1988). La Comunicación Química en Insectos Sociales: Feromonas de Alarma y de Pista. Ecología, N° 2. Icona, Madrir. Pp321-331.
- Hill, P., Wyse, M. & Anderson, D. (2006) Fisiología Animal. Primera Edición. Editorial Médica Panamericana. Madrid, España. Pp.: 609-633.
- Issa, S. (1995) Algunas observaciones sobre el comportamiento de soldados de dos especies de Nasutitermes (Termitidae: Nasutitermitinae). Bol .Entomol. Venez. 10(1):119.
- Moore, B. P., 1968: Studies of the chemical composition and function of the cephalic gland secretion in Australian termites. Inse. Physiol., 14: 33-39.
- Noirot, C. & Noirot-Timothèe, C. (1965) Organisation de la glande sternale chez Calotermes flavicollis F. (Insecta: Isoptera). Étude au microscope électronique. Compte Rendu de l’Académie des Sciencies. 260(23):6202-6204.
- Stuart, A. (1969). Social behavior and communication. Krishna K, Weesner F, Editores. Biology of termites. Academic Press, Inc. New York: USA. Pp: 193-232.
- Pasteels, J. (1965) Polyéthisme chez les ouvriers de N. lujae (Termitidae: Isoptères). Biologia Gabonica 1:191-205.
- Randall, D., Burggren, W. & French, K. (2002) Fisiología Animal: Mecanismos y Adaptaciones. Cuarta Edición. McGraw-Hill Interamericana.Madrid, España. Pp.: 507- 547.
- Resh, V. & R. Cardé. (2009). Encyclopedia of Insects. Editorial Elsevier. Madrid, España. Pp:685-686.
- Traniello, J. (1981) Enemy deterrence in the recruitment strategy of soldier organized foraging in Nasutitermes costalis. Proc. Nat. Acad. Sci. 78(3):1976-1979.
Hola @khrisaeroth El equipo de curación de Proyecto Witness Cervantes encargado de curar contenido técnico y científico considera que tu post es de gran calidad, original, útil y cumple con los requisitos para ser votado. ¡Continúa así!
Muchísimas gracias! y disculpen la demora para contestarles. Abrazos, seguiré el trabajo duro
Congratulations @khrisaeroth! You have completed some achievement on Steemit and have been rewarded with new badge(s) :
Click on any badge to view your own Board of Honor on SteemitBoard.
For more information about SteemitBoard, click here
If you no longer want to receive notifications, reply to this comment with the word
STOP